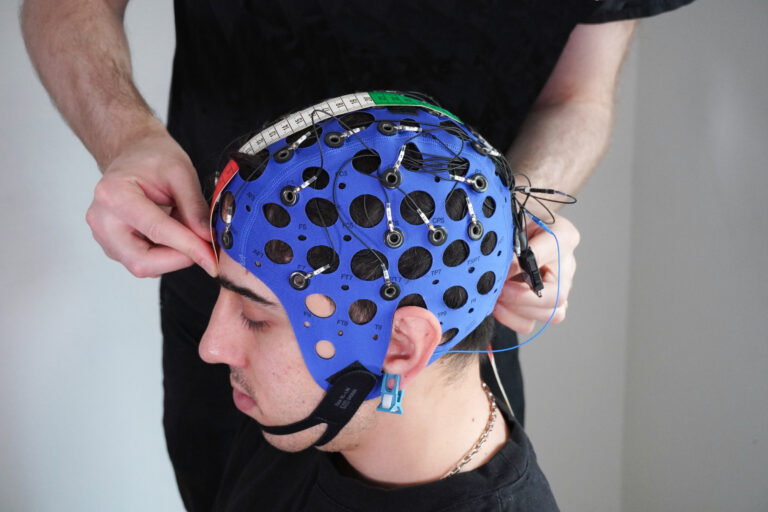
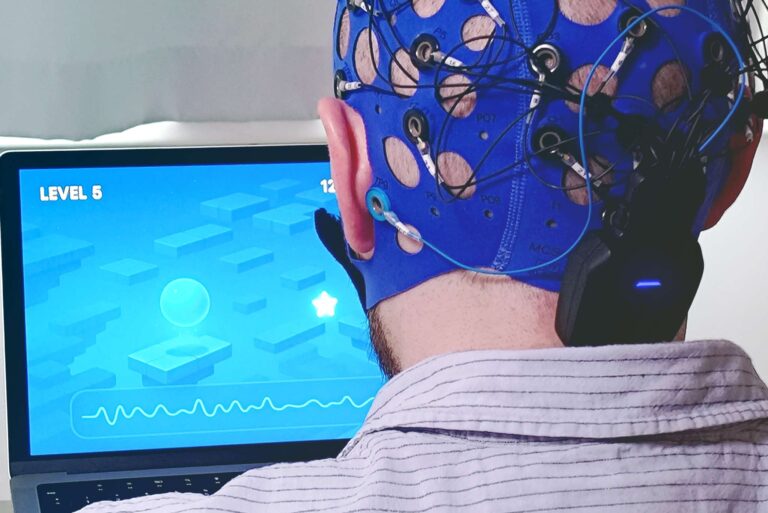
Putting is a key element of golf. A putter is used to strike a ball towards the hole when the ball lies on or just short of the area known as the green.
Understanding the processes underpinning putting expertise is critical. It has been reported that putting accounts for 41% of shots per round of golf (PGA Tour, 2023) making putting performance one of the key factors determining a golf player’s earnings (Alexander & Kern, 2005; Hellstrom, 2009).
Why use EEG to study golf putting performance?
Electroencephalography (EEG) enables us to measure brain activity by capturing changes in electrical potential across the scalp. By interpreting these changes, we can find the brain processes associated with expert sporting performance (Park et al., 2015).

Interested in EEG?
EEG enables us to measure brain wave activity in real time without modifying behavior (Kranczioch et al., 2014; Ojeda et al., 2014). In fact, mobile EEG, like Mentalab Explore, is so small that it does not interfere with putting posture. The golfer does not need to wear a backpack to store the amplifier, nor have excess wires running down their neck and back.
Depending on the exact configuration, Explore amplifiers weigh only 27-36 grams and can be attached to a comfortable neoprene cap that holds the electrodes in place.
High temporal resolution
EEG is excellent at measuring the neural mechanisms associated with sporting performance.
It provides a higher temporal resolution (milliseconds) than other brain imaging techniques, allowing us to measure complex behavior performed over a short period of time, and we can time-lock EEG to the self-paced pre-performance routine of a performer.
This is important. Prior to motor execution, golfers engage in a motor phase pre-shot (“from address to contact” in golf parlance). There are limited movement artifacts during pre-shot routines as golfers’ feet remain still and their motor actions subtle.
One might think the putting action itself would then cause head and eye movement artifacts in the EEG signal. However, our previous research and that of others has not reported any concerns or data loss due to these movement artifacts.
Neural mechanisms associated with golf putting performance
To explore the neural mechanisms associated with golf putting, researchers tend to use power spectral density analysis and time frequency analysis. These methods allow us to explore the changes in brain activity that are related to specific behaviors or times of interest.
For power spectral density analysis and time frequency analysis, a priori frequency bands (based on the wider neuroscience literature and sporting literature) are typically selected. For example, the theta (4-7 Hz), alpha (8-12 Hz), and beta (15-30 Hz) frequency bands, and the sensorimotor rhythm (SMR; 13-15 Hz).
Preparation is key
The superior performance of a highly skilled golfer may be related to their preparation.
In our research, successful putts were associated with increased attention while aiming (theta), and greater and earlier suppression of beta and SMR frequencies during the preparation phase.

Beta and SMR frequencies in successful putts were maximal 2000ms to 1000ms before contact with the ball.
Decreases in beta frequencies may indicate that experts devote more neural resources to the response programming of their putt (Cooke et al., 2014). That is, they may actively program the movement direction and force in the preparation phase (Neubauer & Fink, 2009; Pfurtscheller, 1992).
We also found that successful putts were associated with an increase in relative power (%) from baseline in the alpha frequency at electrode C4. This occurred 1000ms to 500ms before contact.
Changes in relative alpha power at C4 has been linked to enhanced fine motor control (Cheron et al., 2016). This suggests functional inhibition of the left hand and arm during successful putts (Klimesch et al., 2007).
The left arm and hand in golf putting is thought to influence the direction of the putter face and help keep the putter square at contact.
Inconsistency: challenges remain
Findings in the wider literature are inconsistent.
It is unclear whether an increase or decrease in alpha power is related to successful golf putting performances (Babiloni et al., 2008; Baumeister et al., 2008; Cooke et al., 2014) or indeed whether a change in alpha power is linked to putting performance at all (Cooke et al., 2015).
This highlights the complex relationship between baseline alpha power and subsequent cognitive-motor processes.
It also speaks to the methodological challenges which are particularly acute for elite performers. These include changes in hole size, and the difficulty in using “standardised putters” (Babiloni et al., 2008; Cooke et al., 2014).
If a task changes, golfers may become unfamiliar with the task and changes in sensory information and feedback ensue.
Across studies, there has also been inconsistency in the number of electrodes chosen, recording equipment, and choice of analysis, including filtering and baseline. These factors impact replication.
Why Mentalab Explore may be a solution
By allowing golfers to use their own putter on a ZEN Greenstage, and adopting a standardised protocol with Mentalab Explore amplifiers, we hope to create a controlled, experimental design that correctly represents real, competitive putting.
Representing such an environment is essential if we wish to investigate the psychology of competitive golf.
In a representative task design, golfers complete putts from different locations. To succeed, a golfer must adapt to each location.
To properly capture this adaption process, golfers must be allowed to complete their full pre-shot routine, which can be up to 40 seconds in length. They must be provided the opportunity to search for perceptual information in the environment by scanning the green before each putt.

Current research designs tend to be less representative. Here, golfers are presented with a high number of repetitive putts, often taken from one location. This ultimately results in shorter, pre-performance routines – participants tend not to move their feet between putts and scan the green less. In essence, there is no need to account for changes in the environment or continually read the green.
Previously, we compared the results of research that used representative and unrepresentative task designs. We found that the more representative the task, the more accurate the results.
Future work
Examining neural activity may provide insight into the processes that underpin perception and action (Wilson et al., 2016). In turn, we may gain information about the behavioral and neural mechanisms of performance-enhancing strategies that are used by expert performers (Gonzalez et al., 2017).
Critically, we still require longitudinal EEG studies that explore the development of expertise in putting (Park et al., 2015). We will be unable to accurately guide coaches, practitioners and golfers before this work is done.
Mentalab Explore could help
The positive feedback we received from golfers who wore Mentalab Explore highlights how comfortable the system is, and how little it impacts performance. The system’s wearability and quick set up time means it is more feasible to collect longitudinal data in the future.
Additionally, we can decrease set up time further by using the 32-channel system once to create a baseline profile, and then following up with 8-channel systems.
We are excited about the prospect of collecting more research in this vain to inform applied practice.
Interested in measuring brain responses during high performance sport? Let’s have a chat! Reach out at
References
Alexander, D. L., & Kern, W. (2005). Drive for Show and Putt for Dough?: An Analysis of the Earnings of PGA Tour Golfers. Journal of sports economics, 6(1), 46-60. https://doi.org/10.1177/1527002503260797
Babiloni, C., Del Percio, C., Lacoboni, M., Infarinato, F., Lizio, R., Marzano, N., . . . Eusebi, F. (2008). Golf putt outcomes are predicted by sensorimotor cerebral EEG rhythms. Journal of Physiology-London, 586(1), 131-139. https://doi.org/10.1113/jphysiol.2007.141630
Baumeister, J., Reinecke, K., Liesen, H., & Weiss, M. (2008). Cortical activity of skilled performance in a complex sports related motor task. European Journal of Applied Physiology, 104(4), 625-631. https://doi.org/10.1007/s00421-008-0811-x
Cheron, G., Petit, G., Cheron, J., Leroy, A., Cebolla, A., Cevallos, C., . . . Dan, B. (2016). Brain Oscillations in Sport: Toward EEG Biomarkers of Performance. Frontiers in Psychology, 7, Article 246. https://doi.org/10.3389/fpsyg.2016.00246
Cooke, A., Gallicchio, G., Kavussanu, M., Willoughby, A., McIntyre, D., & Ring, C. (2015). Premovement high-alpha power is modulated by previous movement errors: Indirect evidence to endorse high-alpha power as a marker of resource allocation during motor programming. Psychophysiol, 52(7), 977-981. https://doi.org/10.1111/psyp.12414
Cooke, A., Kavussanu, M., Gallicchio, G., Willoughby, A., McIntyre, D., & Ring, C. (2014). Preparation for action: Psychophysiological activity preceding a motor skill as a function of expertise, performance outcome, and psychological pressure. Psychophysiol, 51(4), 374-384. https://doi.org/10.1111/psyp.12182
Gonzalez, C. C., Causer, J., Miall, R. C., Grey, M. J., Humphreys, G., & Williams, A. M. (2017). Identifying the causal mechanisms of the quiet eye. European Journal of Sport Science, 17(1), 74-84. https://doi.org/10.1080/17461391.2015.1075595
Hellstrom, J. (2009). Competitive Elite Golf: A Review of the Relationships between Playing Results, Technique and Physique. Sports Med, 39(9), 723-741. https://doi.org/10.2165/11315200-000000000-00000
Klimesch, W., Sauseng, P., & Hanslmayr, S. (2007). EEG alpha oscillations: The inhibition-timing hypothesis. Brain Research Reviews, 53(1), 63-88. https://doi.org/10.1016/j.brainresrev.2006.06.003
Kranczioch, C., Zich, C., Schierholz, I., & Sterr, A. (2014). Mobile EEG and its potential to promote the theory and application of imagery-based motor rehabilitation: Towards mobile EEG. International journal of psychophysiology, 91(1), 10-15.
Neubauer, A. C., & Fink, A. (2009). Intelligence and neural efficiency: Measures of brain activation versus measures of functional connectivity in the brain. Intelligence (Norwood), 37(2), 223-229. https://doi.org/10.1016/j.intell.2008.10.008
Ojeda, A., Bigdely-Shamlo, N., & Makeig, S. (2014). MoBILAB: an open source toolbox for analysis and visualization of mobile brain/body imaging data. Front Hum Neurosci, 8, 121-121. https://doi.org/10.3389/fnhum.2014.00121
Park, J. L., Fairweather, M. M., & Donaldson, D. I. (2015). Making the case for mobile cognition: EEG and sports performance. Neurosci Biobehav Rev, 52, 117-130. https://doi.org/10.1016/j.neubiorev.2015.02.014
Pfurtscheller, G. (1992). EVENT-RELATED SYNCHRONIZATION (ERS) – AN ELECTROPHYSIOLOGICAL CORRELATE OF CORTICAL AREAS AT REST. Electroencephalography and Clinical Neurophysiology, 83(1), 62-69. https://doi.org/10.1016/0013-4694(92)90133-3
Wilson, M, Wood, G, Vine, S. (2016). Say it quietly, but we still do not know how Quiet Eye training works – comment on Vickers. Current Issues in Sport Science, 1. https://doi.org/10.15203/CISS_2016.117